
Theropoda
Sabías ...
SOS Children hizo esta selección Wikipedia junto a otros recursos de escuelas . Madres SOS cada aspecto después de un una familia de niños apadrinados .
| Los terópodos Rango temporal: Triásico Tardío-presente, 231.4-0Ma | |
|---|---|
 | |
| Montado réplica de un Tyrannosaurus rex esqueleto | |
| Clasificación científica | |
| Reino: | Animalia |
| Filo: | Chordata |
| Clase: | Reptilia |
| Orden: | Saurisquia |
| Clade: | Eusaurischia |
| Clade: | Theropoda Marsh, 1881 |
| Subgrupos | |
| |
Theropoda (terópodo / θ ɛr ə p ɒ d /; Nombre del suborden Theropoda / θ ɨ r ɒ p ɵ d ə /, Del griego y significa "pies bestia") es a la vez un suborden de bípedos saurischian dinosaurios , y un clado que consiste en que suborden y sus descendientes (incluyendo modernas aves ). Los dinosaurios pertenecientes al suborden Theropoda fueron principalmente carnívora , aunque un número de grupos de terópodos evolucionó herbivoría , omnivoría, y insectivoría. Los terópodos aparecieron por primera vez durante el Edad Cárnico de finales del Triásico periodo de hace unos 230 millones de años ( Ma) e incluyó los únicos grandes carnívoros terrestres de la Jurásico temprano por lo menos hasta el cierre del Cretácico , alrededor de 65 Ma. En el Jurásico , las aves evolucionaron de pequeños especializada terópodos coelurosaurian, y son hoy representado por 9.900 especies vivas.
Entre las características que unen los dinosaurios terópodos de aves son los pies de tres dedos, un furcula (horquilla), huesos llenos de aire, melancólico de la huevos, y (en algunos casos) plumas.
Biología
Dieta

Los terópodos exhiben una amplia gama de dietas, de los insectívoros a los herbívoros y carnívoros. Carnivory Estricto siempre ha sido considerada como la dieta ancestral de los terópodos, como grupo, y una variedad más amplia de las dietas fue históricamente considerada una característica exclusiva de los terópodos aviares (aves). Sin embargo, los descubrimientos en los finales del siglo 20 y principios de los 21 demostraron que una variedad de dietas existía incluso en más linajes basales. Todos los hallazgos iniciales de fósiles de terópodos mostraron que sean principalmente carnívora . Especímenes fosilizados de los primeros terópodos conocidos por los científicos en los siglos 19 y 20 todos poseían dientes afilados con bordes dentados para cortar carne, y algunos ejemplares incluso mostraron pruebas directas de la práctica abusiva. Por ejemplo, un Compsognathus longipes fósil fue encontrado con un lagarto en su estómago, y un Velociraptor mongoliensis espécimen se encontró encerrado en combate con un andrewsi Protoceratops (un tipo de ornitisquio dinosaurio).
Los terópodos fósiles no carnívoras primera confirmados encontradas fueron la therizinosaurs, originalmente conocidos como segnosáuridos. Primero piensa que es prosaurópodos, estos dinosaurios enigmáticos fueron posteriormente demostrado ser altamente especializados herbívoros terópodos. Therizinosaurs poseían grandes abdómenes para la elaboración de alimentos de las plantas, y las pequeñas cabezas con picos y dientes en forma de hoja. Continuando el estudio de terópodos maniraptoran y sus relaciones mostraron que therizinosaurs no eran los únicos miembros tempranos de este grupo a abandonar carnivory. Varios otros linajes de los primeros maniraptors muestran adaptaciones para un dieta omnívora, incluidas las semillas que comen (algunos troodontids) y come insectos (muchos avialans y alvarezsaurs). Oviraptorosaurs, ornitomimosaurios y troodontids avanzadas probablemente omnívora, así, y algunos terópodos tempranas (como Masiakasaurus knopfleri y la espinosáuridos) parecen haberse especializado en la captura de peces.
Piel, escamas y plumas
Terópodos Mesozoico eran también muy diversa en términos de textura de la piel y la cubierta. Plumas o estructuras similares a plumas son atestiguados en la mayoría de los linajes de los terópodos. (Ver dinosaurio emplumado). Sin embargo, fuera de la coelurosaurs, plumas pueden haber sido confinado a los jóvenes, las especies más pequeñas, o partes del animal limitado. Muchos terópodos más grandes habían piel cubierta de escamas pequeñas, llenas de baches. En algunas especies, éstas se intercalan con las escalas más grandes con núcleos óseos u osteodermos. Este tipo de piel es el más conocido en el ceratosauro Carnotaurus, que se ha conservado con extensas impresiones de piel.
A partir de 2012, el terópodo protofeathered conocida más divergente es Sciurumimus. Por lo tanto, se puede inferir que Tetanurae eran al menos parcialmente con plumas; sin evidencia de (proto) plumas se ha descubierto entre la Ceratosauria. Los linajes más distantes de las aves tenían plumas que eran relativamente corto y compuesto por filamentos simples, posiblemente de ramificación. Filamentos simples también se observan en therizinosaurs, que también poseían grandes, endurecieron "pluma" -como plumas. Más plenamente terópodos con plumas, como dromaeosaurios, por lo general conservan escalas sólo en los pies. Algunas especies pueden tener plumas mixtos otras partes del cuerpo también. Scansoriopteryx conserva escalas cerca de la parte inferior de la cola, y Juravenator puede haber sido predominantemente escamosa con algunos filamentos simples intercalados. Por otro lado, algunos terópodos estaban completamente cubiertos con plumas, como el troodóntido Anchiornis, que incluso tenía plumas en los pies y los dedos de los pies.
Tamaño
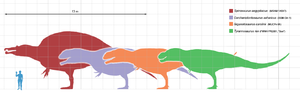
Tyrannosaurus era la más grande y popular terópodo conocido por el público en general durante muchas décadas. Desde su descubrimiento, sin embargo, una serie de otros dinosaurios carnívoros gigantes se han descrito, incluyendo Spinosaurus , Carcharodontosaurus y Giganotosaurus . Los ejemplares originales Spinosaurus (así como los fósiles más recientes descritos en 2006) apoyan la idea de que el Spinosaurus es más grande que el Tyrannosaurus, demostrando que Spinosaurus era posiblemente 6 metros más largo y al menos 1 tonelada métrica más pesado que el Tyrannosaurus. Todavía no hay una explicación clara para exactamente por qué estos animales crecían mucho más grande que los depredadores de la tierra que vinieron antes y después de ellos.
El terópodo no avialan más pequeño conocido de ejemplares adultos es la troodóntido Anchiornis huxleyi, en 110 gramos de peso y 34 centímetros (1 pie) de longitud. Cuando se incluyen las aves modernas, el Zunzuncito Mellisuga helenae es más pequeña en el 1,9 gy 5,5 cm (2,2 pulgadas) de largo.
Postura y la marcha
Como un grupo muy diverso de animales, la postura adoptada por terópodos probable variaba considerablemente entre los diferentes linajes a través del tiempo. Todos los terópodos conocidos son bípedo, con las extremidades anteriores reducidos en longitud y especializado para una amplia variedad de tareas (véase más adelante). En las aves modernas, el cuerpo se lleva a cabo típicamente en una posición algo vertical, con la pierna superior (fémur) mantenida en paralelo a la columna vertebral y con la fuerza de avance de la locomoción generada en la rodilla. Los científicos no están seguros de qué tan atrás en el árbol familiar terópodo este tipo de postura y la locomoción se extiende.
Terópodos no aviares fueron reconocidos por primera vez como bípedo durante el siglo 19, antes de su relación con las aves fue ampliamente aceptada. Durante este período, tales como terópodos carnosaurios y se pensaba que los tiranosáuridos haber caminado con fémures verticales y espinas en una posición vertical, casi postura erguida, con su cola larga y musculares como apoyo adicional en una postura tripodal-canguro similares. A partir de la década de 1970, los estudios biomecánicos de los terópodos gigantes extintos ponen en duda esta interpretación. Estudios de articulación ósea de las extremidades y la relativa ausencia de pruebas de huellas de arrastre cola sugirieron que, al caminar, el gigante, terópodos cola larga habría adoptado una postura más horizontal con la cola en paralelo al suelo. Sin embargo, la orientación de las piernas en estas especies al caminar sigue siendo controvertido. Algunos estudios apoyan un fémur orientada verticalmente tradicional, al menos en las grandes terópodos de cola larga, mientras que otros sugieren que la rodilla se flexiona normalmente fuertemente en todos los terópodos al caminar, incluso gigantes como los tiranosáuridos. Es probable que una amplia gama de posturas corporales, posturas, y aires existía en los muchos grupos de terópodos extintos.
Sistema nervioso y sentidos
Aunque raras yesos, completos de terópodo endocrania son conocidos a partir de fósiles. Terópodo endocrania también puede reconstruirse a partir de los casos cerebrales conservados sin dañar ejemplares valiosos utilizando una tomografía computarizada y software de reconstrucción 3D. Estos hallazgos son de importancia evolutiva porque ayudan a documentar la aparición de la neurología de las aves modernas de la de reptiles anteriores. Un aumento en la proporción del cerebro ocupado por el cerebro parece haber ocurrido con el advenimiento de la Coelurosauria y "continuado a lo largo de la evolución de manirraptores y madrugadores ".
Morfología Miembro Anterior

Patas delanteras más cortas en relación con las patas traseras era un rasgo común entre los terópodos, sobre todo en el abelisáuridos (tales como Carnotaurus) y el tiranosáuridos (como Tyrannosaurus ). Este rasgo fue, sin embargo, no es universal: espinosáuridos habían extremidades anteriores muy desarrollado, por lo que también lo hicieron muchos coelurosaurs. Un género, Xuanhanosaurus, también ha afirmado haber sido cuadrúpedo debido a sus relativamente robustas patas delanteras, pero esto ya no piensa que es probable.
Las manos también son muy diferentes entre los distintos grupos; la más común es un apéndice que consta de 3 dedos, los dígitos I, II y III (o, posiblemente, II, III y IV) con garras afiladas. Algunos terópodos basales ( Herrerasaurus , Eoraptor) tenía 4 dígitos, y también un reducido V. metacarpiano Ceratosaurians normalmente tenía 4 dígitos, mientras que la mayoría tetanurans tenían 3.
Ámbito de aplicación de uso Los miembros anteriores también se cree que también han sido diferentes entre diferentes familias. La espinosáuridos podrían haber utilizado sus poderosas patas delanteras para mantener el pescado. Algunos pequeños coelurosaurus ( dromaeosáuridos, scansoriopterygids) se cree que han utilizado sus patas delanteras para subir en los árboles, y las aves, que se cree que son descendientes de los terópodos, los utilizan para el vuelo.
Movimiento Miembro Anterior

En contra de la forma en que los terópodos menudo han sido reconstruidas en el arte y los medios de comunicación populares, el rango de movimiento de las extremidades anteriores terópodos era muy limitado, sobre todo en comparación con la destreza de la extremidad anterior de los seres humanos y otros primates. Más notablemente, terópodos y otros dinosaurios saurisquios bípedos (incluidos los bípedos prosauropodos ) no podía pronación sus manos, es decir, no podían girar el antebrazo de manera que las palmas se enfrentan el suelo o hacia atrás hacia las piernas. En los seres humanos, la pronación se consigue por el movimiento de la radio con relación a la cúbito (los dos huesos del antebrazo). En dinosaurios saurischian, sin embargo, el extremo del radio cerca del codo en realidad estaba bloqueado en una ranura del cúbito, impidiendo cualquier movimiento. Movimiento en la muñeca también fue limitado en muchas especies, obligando a todo el antebrazo y la mano de moverse como una sola unidad con poca flexibilidad. En terópodos y prosaurópodos, el único camino para la palma de la mano para hacer frente a la tierra hubiera sido por ensanchamiento lateral de toda la extremidad anterior, como en un pájaro elevando su ala.
En carnosaurios como Acrocanthosaurus, la propia mano retiene un grado relativamente alto de flexibilidad, con los dedos móviles. Este también fue el caso de más terópodos basales como herrerasaurs y dilofosaurios. Coelurosaurios mostró un cambio en el uso del antebrazo, con una mayor flexibilidad en el hombro permitiendo que el brazo para ser levantado hacia el plano horizontal, y aún mayores grados en las aves que vuelan. Sin embargo, en coelurosaurs tales como ornitomimosaurios y especialmente dromaeosaurios, la propia mano había perdido la mayor flexibilidad, con dedos muy inflexibles. Dromaeosaurios y otra manirraptores también mostraron una mayor movilidad en la muñeca no se ve en otros terópodos, gracias a la presencia de un hueso de la muñeca en forma de media luna especializado (el carpo semi-semilunar) que permitió que toda la mano para doblar hacia atrás hacia el antebrazo en la forma de la moderna aves.
Paleopatología
En el año 2001, Ralph E. Molnar publicó un estudio de patologías en terópodo hueso de dinosaurio. Encontró características patológicas en 21 géneros de 10 familias . Patologías se encontraron en terópodos de todo el tamaño del cuerpo aunque eran menos comunes en fósiles de pequeños terópodos, aunque esto puede ser un artefacto de preservación. Son muy ampliamente representadas en las diferentes partes de la anatomía terópodo. Los sitios más comunes de lesiones conservada y enfermedad en dinosaurio terópodo en el costillas y vértebras de la cola. A pesar de ser abundante en las costillas y las vértebras, las lesiones parecen estar "ausente ... o muy rara" en peso primaria huesos que sostienen los cuerpos como el sacro, fémur, y tibia. La falta de lesiones conservados de estos huesos sugiere que fueron seleccionados por la evolución de la resistencia a la rotura. Los sitios menos comunes de lesiones en conserva son los cráneo y extremidades anteriores, con lesiones que ocurren en aproximadamente la misma frecuencia en cada sitio. La mayoría de las patologías que se conservan en los fósiles de terópodos son los restos de lesiones como fracturas, cavidades y perforaciones, a menudo probablemente originario de picaduras. Algunos paleopatologías terópodos parecen ser evidencia de las infecciones , que tendían a ser confinado sólo a pequeñas regiones del cuerpo del animal. La evidencia de malformities congénitas también se han encontrado en los restos de terópodos. Estos descubrimientos pueden proporcionar información útil para la comprensión de la historia evolutiva de los procesos de desarrollo biológico. Insólito fusiones en elementos craneales o asimetrías en la misma son probablemente evidencia de que se está examinando los fósiles de un muy viejo individuo en lugar de una enferma.
Historia evolutiva

Durante el final del Triásico , varios proto-terópodos y terópodos dinosaurios primitivos existía y evolucionó junto al otro.
La primera y más primitiva de los dinosaurios terópodos fueron los carnívoros Eodromaeus y la herrerasaurids de Argentina (así como, posiblemente, el omnívora Eoraptor). Los herrerasaurs existían a principios del Triásico tardío (finales Carniano a Early Norian). Fueron encontrados en América del Norte y América del Sur y, posiblemente, también la India y África del Sur. Los herrerasaurs se caracterizaron por una mosaico de rasgos primitivos y avanzados. Algunos paleontólogos han considerado en el pasado los herrerasaurians ser miembros de Theropoda, mientras que otros teorizaron que el grupo sea saurisquios basales, e incluso pueden haber evolucionado antes de la escisión saurischian ornitisquio. Análisis cladístico tras el descubrimiento de Tawa, otro dinosaurio del Triásico, sugiere los herrerasaurs probables eran los primeros terópodos.
Los terópodos inequívocas primeros y más primitivos (o, alternativamente, "Eutheropoda" - 'terópodos Verdaderos') son los Coelophysoidea. El Coelophysoidea eran un grupo de animales de amplia distribución, ligeramente construidas y potencialmente gregarios. Incluyeron pequeños cazadores como Coelophysis y depredadores (posiblemente) más grandes como Dilophosaurus . Estos animales éxito continuado de la tarde Cárnico (a principios del Triásico Superior) a través de la Toarciense (finales Jurásico Temprano). Aunque en los primeros clasificaciones cladísticos que se incluyeron en el marco del Ceratosauria y considerado una rama lateral de terópodos más avanzados, que pueden haber sido ancestral a todos los otros terópodos (lo que los haría un grupo parafilético).
El algo más avanzado ceratosaurios (incluidos Ceratosaurus y Carnotaurus) apareció durante el jurásico temprano y continuó hasta el Jurásico Tardío en Laurasia. Ellos compitieron junto a sus más anatómicamente avanzada parientes tetanuros y en la forma de la Abelisauroidea linaje duró-al final del Cretácico en Gondwana .
La Tetanurae son más especializado de nuevo que los ceratosaurios. Se subdividen en la basal Megalosauroidea (alternativamente Spinosauroidea) y el derivado más Avetheropoda. Megalosauridae eran principalmente Jurásico Medio a principios del Cretácico depredadores, y su restos parientes espinosáurido 'son en su mayoría de las rocas del Cretácico Inferior y Medio. Avetheropoda, como su nombre lo indica, fueron más estrechamente relacionados con las aves y se dividen de nuevo en el Allosauroidea (la diversa carcharodontosaurs) y la Coelurosauria (un grupo de dinosaurios muy grande y diversa, incluyendo las aves).
Así, durante el Jurásico tardío, había no menos de cuatro linajes distintos de terópodos-ceratosaurios, megalosaurs, alosaurios y coelurosaurs-se aprovechan de la abundancia de pequeñas y grandes dinosaurios herbívoros. Los cuatro grupos sobrevivieron hasta el Cretácico, y tres de ellos-el ceratosaurios, coelurosaurs y alosaurios-sobrevivieron al final del período, donde estaban geográficamente aislada, los ceratosaurios y alosaurios en Gondwana, y los coelurosaurs en Laurasia.
De todos los grupos de terópodos, los coelurosaurs fueron, con mucho, el más diverso. Algunos grupos celurosaurio que florecieron durante el Cretácico eran la tiranosáuridos (incluyendo Tyrannosaurus ) la dromaeosáuridos (incluyendo Velociraptor y Deinonychus , que son muy similares en forma a las más antiguas conocidas de aves, Archaeopteryx ), el pájaro-como troodontids y oviraptorosaurs, la ornitomimosaurios (o "dinosaurios avestruz"), el extraño gigante garra herbívoro therizinosaurs, y el avialans, que incluyen las aves modernas y es el único linaje de los dinosaurios para sobrevivir a la Cretácico-Paleógeno evento de extinción. Si bien las raíces de estos diversos grupos se encuentran en el Jurásico Medio, sólo fueron abundantes durante el Cretácico Temprano. Unos paleontólogos, tales como Gregory S. Paul, han sugerido que algunos o todos estos terópodos avanzados eran en realidad descendientes de los dinosaurios voladores o proto-aves como Archaeopteryx que perdieron la capacidad de volar y devueltos a un hábitat terrestre.
Clasificación
Historia de la clasificación

El nombre Theropoda (que significa "pies bestia") fue acuñado por primera vez por OC Marsh en 1881. Marsh nombró inicialmente Theropoda como suborden para incluir el familia Allosauridae, pero más tarde amplió su ámbito de aplicación, volver a lo que la sitúa como una para incluir una amplia gama de "carnívoros" familias de dinosaurios, incluyendo Megalosauridae, Compsognathidae, Ornithomimidae, Plateosauridae y Anchisauridae (ahora conocido por ser herbívoro prosauropodos) y Hallopodidae (ahora conocido por ser parientes de los cocodrilos). Debido al alcance de la de Marsh Orden Theropoda, que vino a sustituir a un grupo taxonómico anterior que el rival de Marsh ED Cope había creado en 1866 por los dinosaurios carnívoros, Goniopoda ("pies en ángulo").
A principios del siglo 20, algunos paleontólogos, como Friedrich von Huene, ya no se considera dinosaurios carnívoros que han formado un grupo natural. Huene abandonó el nombre Theropoda, en lugar de utilizar Harry Seeley Orden Saurisquia , que Huene divide en los subórdenes Coelurosauria y Pachypodosauria . Huene coloca la mayor parte de los pequeños grupos de terópodos en Coelurosauria, y los grandes terópodos y prosaurópodos en Pachypodosauria, que consideraba a la ancestral Sauropoda (prosaurópodos todavía eran considerados como carnívoros en este momento, debido a la asociación incorrecta de cráneos y dientes con cuerpos prosaurópodo rauisuchian, en animales tales como Teratosaurus). En WD Mateo y De Barnum Brown 1922 descripción de la primera conocida dromeosáurido ( Dromaeosaurus albertensis), se convirtieron en los primeros paleontólogos para excluir prosauropodos de los dinosaurios carnívoros, y trató de revivir el nombre Goniopoda para ese grupo, aunque ninguna de estas sugerencias fueron aceptadas por otros científicos.

No fue sino hasta 1956 que Theropoda volvió a entrar en su uso como taxón que contiene los dinosaurios carnívoros y sus descendientes, cuando Alfred Romer reclasificado la Orden Saurisquia en dos subórdenes, Theropoda y Sauropoda. Esta división básica ha sobrevivido en la paleontología moderna, con la excepción de, de nuevo, el Prosauropoda, que incluye como Romer infraorden de los terópodos. Romer también mantuvo una división entre Coelurosauria y Carnosauria (que él también se clasificó como infraórdenes). Esta dicotomía fue derrotado por el descubrimiento de Deinonychus y Deinocheirus en 1969, ninguno de los cuales podría clasificarse fácilmente como "carnosaurios" o "coelurosaurs." A la luz de estos y otros descubrimientos, a finales de 1970 Rinchen Barsbold creado una nueva serie de infraórdenes terópodos: Coelurosauria, Deinonychosauria, Oviraptorosauria, Carnosauria, Ornithomimosauria, y Deinocheirosauria.
Con el advenimiento de cladística y nomenclatura filogenética en la década de 1980, y su desarrollo en los años 1990 y 2000, una imagen más clara de las relaciones terópodos comenzaron a surgir. Varios grupos grandes terópodos fueron nombrados por Jacques Gauthier en 1986, incluyendo la clado Tetanurae por una rama de un terópodo básica dividida con otro grupo, la Ceratosauria. Como más información acerca de la relación entre los dinosaurios y las aves salió a la luz, los más terópodos similares a las aves fueron agrupados en el clado Maniraptora (también llamado por Gauthier en 1986). Estos nuevos desarrollos también vino con un reconocimiento entre la mayoría de los científicos que las aves surgieron directamente de los terópodos maniraptoran y, con el abandono de las filas en la clasificación cladístico, la reevaluación de las aves como un subconjunto de los dinosaurios terópodos que sucedió haber sobrevivido a las extinciones del Mesozoico en el presente.
Los grupos principales



La siguiente es una clasificación simplificada de los grupos de terópodos en función de sus relaciones evolutivas, y organizados en base a la lista de especies de dinosaurios mesozoicos proporcionados por Holtz (2008). Una versión más detallada se puede encontrar en Clasificación de los dinosaurios. La cruz (†) se utiliza para significar grupos sin miembros vivos.
- † Herrerasauria (primeros carnívoros bípedos)
- † Coelophysoidea (pequeños, los primeros terópodos; incluye Coelophysis y otros familiares cercanos)
- † Dilophosauridae (a principios de cresta y terópodos carnívoros)
- † Ceratosauria (generalmente elaborados con cuernos, los carnívoros meridionales dominantes del Cretácico)
- Tetanurae ("colas rígidas"; incluye la mayoría de los terópodos)
- † Megalosauroidea (grupo a principios de los grandes carnívoros, incluyendo los espinosáuridos semiacuáticos)
- † Carnosauria ( Allosaurus y otros familiares cercanos, como Carcharodontosaurus)
- Coelurosauria (eran terópodos, con una gama de tamaños de cuerpo y nichos)
- † Compsognathidae (coelurosaurs principios comunes con los miembros delanteros reducidos)
- † Tyrannosauridae ( Tyrannosaurus y parientes cercanos; había reducido extremidades anteriores)
- † Ornithomimosauria (" avestruz -mimics "; en su mayoría sin dientes; carnívoros a herbívoros posibles)
- † Alvarezsauroidea (pequeños insectívoros con extremidades delanteras reducidas cada una con una garra ampliada)
- Maniraptora ("ladrones de la mano"; tenían brazos largos y delgados y dedos)
- † Therizinosauria (herbívoros bípedos con grandes garras de la mano y la cabeza pequeña)
- † Oviraptorosauria (en su mayoría sin dientes; su dieta y estilo de vida son inciertos)
- † Archaeopterygidae (pequeño, terópodos alados o aves primitivas)
- † Deinonychosauria (pequeñas y medianas empresas; de pájaro, con una garra distintiva dedo del pie)
- Avialae (aves modernas y parientes extintos)
- † Scansoriopterygidae (pequeños avialans primitivos con terceros dedos largos)
- † Omnivoropterygidae (grande, avialans corto cola primeros)
- † Confuciusornithidae (pequeños avialans sin dientes)
- † Enantiornithes (primitivo árbol-vivienda, avialans voladores)
- Euornithes (aves voladoras avanzada)
- † Yanornithiformes (dentado pájaros chinos Cretácico)
- † Hesperornithiformes (aves acuáticas de buceo especializados)
- Aves (, aves picudas modernos y sus parientes extintos)
Relaciones
La siguiente cladograma es una adaptación de Weishampel et al., 2004. Conserva Coelophysoidea como sea posible ceratosaurios, a diferencia de muchos estudios recientes colocándolos fuera ceratosaurios y ancestral tanto ceratosaurios y Tetanurae.
| Theropoda |
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
